


膀胱过度活动症—OAB调控机制研究进展

膀胱是一个典型的躯体神经和植物神经双重支配的储尿/排尿器官,其兴奋性是指调节膀胱作为一个整体器官,执行储尿和排尿的能力。 在膀胱统一协调的兴奋和收缩活动中,神经系统、膀胱上皮、平滑肌层等多个组织和细胞均发挥作用。 膀胱兴奋性异常是导致膀胱过度活动(OAB)(注:膀胱过度活动与间质性膀胱炎的症状有相似,两者诊断容易混淆,造成误诊)产生的主要原因,长期以来,关于OAB的发生机制一直存在较大争议,神经源性学说和肌源性学说均从不同侧面解释了OAB发生的机制,但两者均存在各自的优势和局限性,无法完全自圆其说。
而事实上,越来越多的研究证据也表明,OAB的发生涉及的机制十分复杂,神经元、上皮细胞、平滑肌细胞、间质细胞等因素导致的膀胱兴奋性异常均在OAB的发生中扮演者不一样的角色。
其主要涉及兴奋性调控机制有四个方面:
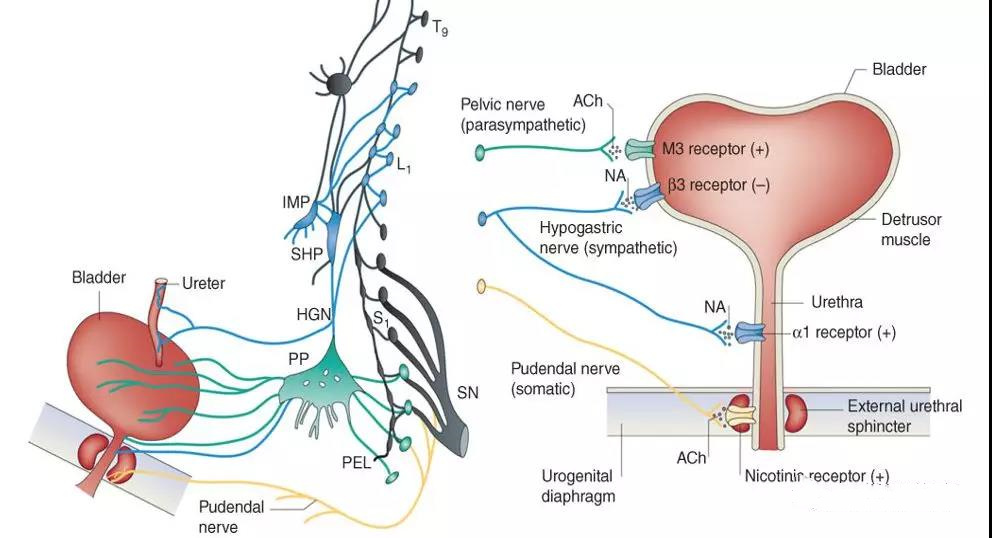
1. 膀胱上皮细胞在膀胱兴奋性调控的作用
一直以来,膀胱上皮层被认为仅仅在膀胱兴奋调节中起“屏障”作用,主要是隔离尿液及其下层的膀胱组织。最新的研究表明,膀胱上皮在膀胱感觉传导和兴奋性调控中发挥着“感觉传导器”的功能,与神经元的作用机制类似。膀胱上皮的完整性是膀胱自发性收缩的重要基础,上皮层的异常可导致膀胱感觉传导和兴奋性的变化。膀胱上皮细胞对兴奋的调控主要是通过自身表达的受体和离子通道的活性调节,以及通过释放活性物质或与其他细胞间相互作用来完成的。
目前的研究认为膀胱上皮表达多种功能受体和离子通道,参与膀胱功能兴奋性的调节。在机械刺激(如膀胱壁张力改变)和化学刺激(如细胞外离子改变)作用下,上皮细胞的受体和离子通道激活,导致多种神经递质如乙酰胆碱(Ach),三磷酸腺苷(ATP)和一氧化氮(NO)等通过释放进入组织间隙,作用于其下层的神经末梢、平滑肌细胞和间质细胞,共同参与了对膀胱感觉传导和兴奋性的调控。
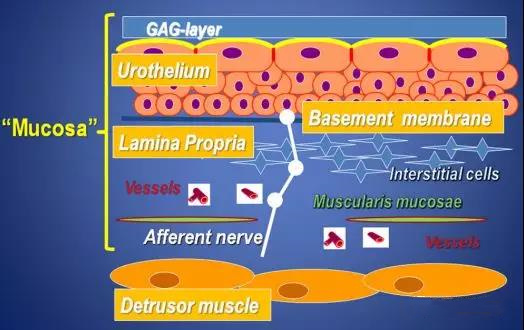
2. 膀胱平滑肌细胞和膀胱兴奋性的关系
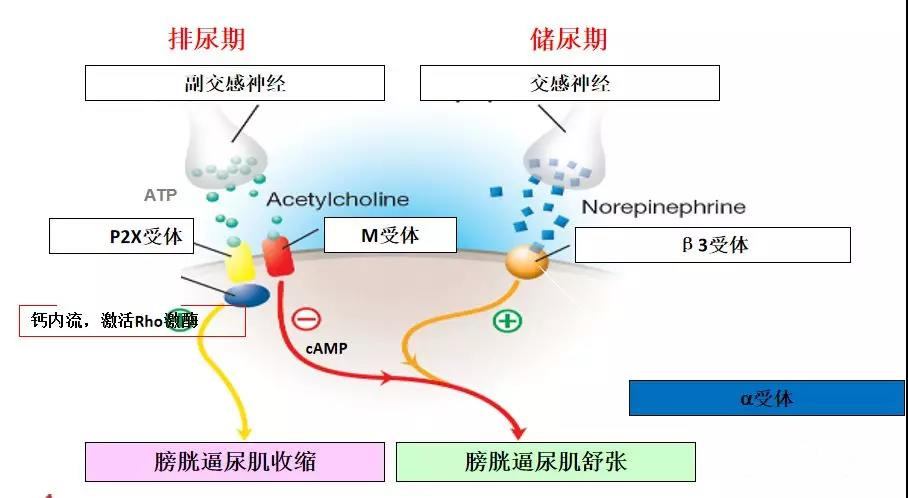
膀胱的排尿和储尿功能有赖于逼尿肌的收缩和舒张,逼尿肌复杂而精细的舒缩功能的调节是由胆碱能神经、肾上腺素能神经、非肾上腺素能非胆碱能神经等共同参与完成的。其中,调节逼尿肌收缩功能的受体主要有胆碱受体和嘌呤受体,调节其舒张功能的受体主要为肾上腺素受体。
胆碱能M受体在逼尿肌中含量丰富,尤以膀胱体部含量最高,膀胱颈和近端尿道平滑肌中亦有少量分布,其兴奋是逼尿肌收缩的主要驱动因素。已证明逼尿肌的主要M受体亚型为M2和M3。M2受体数量较多,但它在正常膀胱组织中不直接介导逼尿肌的收缩,而是通过逆转β肾上腺素能神经介导的逼尿肌舒张来间接介导逼尿肌收缩。M3受体数量较少,但在正常膀胱的收缩活动中起主要作用。临床上,索利那新等选择性M3受体拮抗剂,仍然是膀OAB的主要治疗选择,针对膀胱胆碱能系统开发选择性更高的药物,依然是未来OAB治疗的主要策略方向。
肾上腺素β受体是调节膀胱逼尿肌舒张的主要因素,β受体激活后可以使G蛋白Gs异三聚体解离,激活腺苷酸环化酶,使ATP降解为cAMP,cAMP作为第2信使可激活蛋白激酶A(PKA)产生一系列生物学效应,如钾离子通道开放,肌细胞膜超级化及L型钙离子通道关闭等,从而使松弛逼尿肌。目前研究认为肾上腺素能β3受体是调节大鼠和人逼尿肌舒张的主要亚型。β1受体激动剂(多巴酚丁胺)和β2受体激动剂(丙卡特罗)均不能使人离体逼尿肌明显舒张,而米拉贝隆,作为一种成熟使用的选择性β3受体激动剂,目前被认为是膀胱过度活动症的一线治疗药物之一。
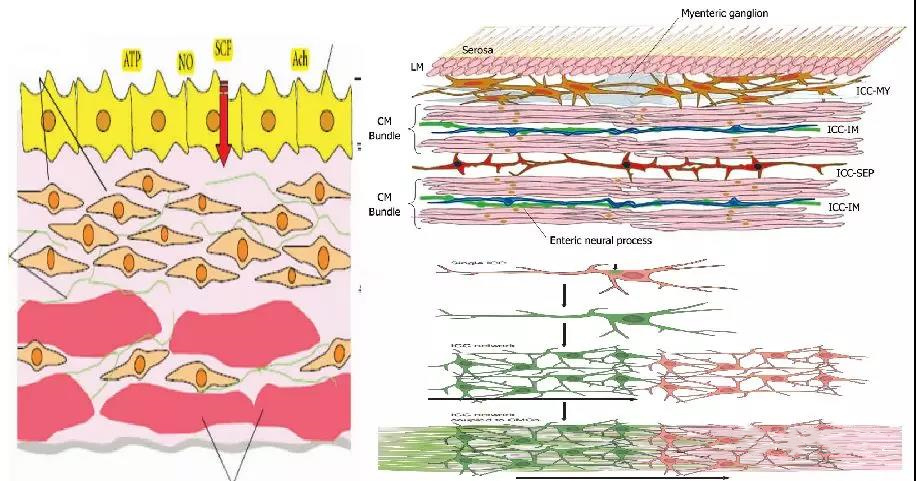
3. ICC样起搏细胞对膀胱兴奋性的调控
1996年,Smet等首次报道了在豚鼠和人逼尿肌上存在类似于胃肠道Cajal间质细胞(ICC)的一种cGMP阳性的特殊细胞,之后的研究进一步证明,这种特殊的细胞还同时表达Vimentin、c-kit、和Ano1等多种分子,特别c-kit一直以来被认为是该细胞的特异性分子标记。之后针对该细胞的研究发现其与胃肠道ICC细胞具有相似的生物学标志,细胞形态和功能特性,遂有研究者将该细胞正式命名为膀胱ICC细胞。现在的观点认为,膀胱中的ICC细胞有四种类型,固有膜ICC,逼尿肌ICC,肌束间星形ICC和血管周ICC。其中,固有膜ICC细胞和逼尿肌内ICC细胞可能是主要的功能亚型,虽然两者都展示出了自发性的细胞内钙离子变化,但是在膀胱兴奋性调控中的作用却不尽相同。
固有膜中的ICC细胞主要存在于上皮下的固有膜中,细胞之间通过缝隙连接CX43蛋白相互连接成网状。现有观点认为,固有膜ICC细胞形成的细胞网络,主要的功能是参与了上皮细胞、神经纤维和平滑肌细胞之间的信号传导。
逼尿肌ICC细胞相互之间没有直接连接,而是散在排布在平滑肌束边缘。目前的观点认为,逼尿肌ICC细胞可能主要是产生肌源性兴奋性和参与对平滑肌兴奋性的微调节。
4. 细胞间通讯
和心肌类似,膀胱逼尿肌也有功能合胞体的特性,正常情况下,逼尿肌在膀胱排空过程中能够实现协调、规律的收缩,其功能调节是由多种因素共同参与的结果。目前大多数人的观点认为正常排尿主要是由神经触发协调一致的逼尿肌收缩引起的。由于逼尿肌细胞和支配其的神经突触并不是“点对点”的结构,因此神经信号的传递并不是同时传导至每个逼尿肌细胞,而排尿收缩需要逼尿肌收缩同步和叠加,因此细胞之间存在一种耦联,使收缩力或电信号由神经支配的肌细胞向无神经支配的肌细胞传递,即细胞耦联。由于正常逼尿肌电传导连接较少,电生理证据显示逼尿肌细胞阈下电流的细胞间传递仅能达到少量细胞[29]。但在某些病理状态下,逼尿肌的电生理特性可随着病理结构改变而发生改变,细胞间电传导也随之出现各种变化。
膀胱兴奋性是维持膀胱功能状态的基础,其调控机制是个复杂而精细的过程。虽然神经在膀胱的兴奋性调节具有主导地位,但是其功能的发挥并不仅仅是“点对点”式的靶向调节。膀胱中上皮细胞、间质细胞、平滑肌细胞甚至炎症细胞等多种组织结构也同样在其中发挥着精细调节的作用,特别是膀胱上皮细胞和ICC细胞的功能角色越来越受到研究者的青睐,两者在膀胱功能调节中的作用也不断被发现和更新,相信进一步的研究会给我们带来更多的惊喜,为膀胱功能障碍性疾病的治疗提供更新的思路和策略。
注释:陆军军医大学附属新桥医院泌尿外科,一直从事尿控相关基础研究,科室李龙坤主任曾就膀胱兴奋调控机制进行了系统综述,文章发表在《中华泌尿外科杂志》。
